4. El lienzo viviente: genética, evolución y estética en la cría del guppy (Poecilia reticulata)
Un análisis de la complejidad hereditaria detrás de los patrones Snakeskin, Tuxedo y la policromía natural
Autor: Javier Rodríguez Luna
Fecha: 29 de Mayo del año 2026
Introducción
El guppy (Poecilia reticulata) ha sido considerado durante décadas uno de los organismos modelo más fascinantes tanto para la biología evolutiva como para la acuariofilia especializada. Su asombrosa plasticidad genética y su rápido ciclo reproductivo permiten observar fenómenos evolutivos y de herencia en tiempo récord. Lo que para un criador aficionado comienza como la búsqueda de un pez estéticamente atractivo, para la ciencia representa un complejo entramado de interacciones alélicas, dominancia incompleta, epistasis y selección artificial. Este artículo analiza los hitos genéticos que revolucionaron la cría de esta especie a partir de la segunda mitad del siglo XX, centrándose en el desarrollo del patrón Snakeskin (piel de serpiente), la fisiología de la expresión cromática y la aparición del fenotipo Halfblack o Tuxedo. A través de estos casos, se explora la tensión constante entre la intervención humana para fijar caracteres estéticos y los límites biológicos impuestos por la herencia ancestral del pez.
#GeneticaAcuatica #PoeciliaReticulata #DivulgacionCientifica #Acuariofilia #EvolucionBiolgica
El enigma del patrón Snakeskin (piel de serpiente)
El desarrollo del patrón Snakeskin representa uno de los logros más significativos en la historia de la selección artificial del guppy. Este diseño, caracterizado por una compleja red de líneas y puntos que imitan la piel de un reptil, no surgió de manera espontánea como una mutación única y limpia, sino como el resultado de un largo proceso de acumulación y ordenamiento de genes modificadores a partir de variaciones ya presentes en las poblaciones silvestres.


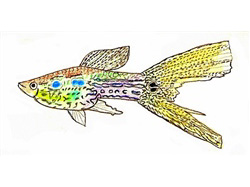
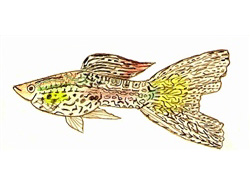
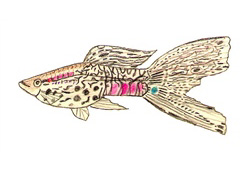

Historia y evolución de una filigrana genética
Las primeras manifestaciones documentadas de este carácter se remontan a finales de la década de 1950, cuando se identificaron los llamados lace-tails (colas de encaje) en ejemplares silvestres. Estos peces exhibían un patrón reticulado fino limitado exclusivamente a la aleta caudal.
[Imagen: Comparativa de un guppy silvestre de "cola de encaje" frente a un ejemplar estabilizado con patrón Snakeskin corporal completo]
Tuvieron que transcurrir casi dos décadas de cría selectiva para que este patrón incompleto se extendiera de forma homogénea a lo largo de todo el cuerpo del pez, culminando en la famosa presentación de esta variedad en la exposición de Manchester en 1966. Este hito no fue el esfuerzo aislado de un solo criador, sino un fenómeno de convergencia en la selección artificial, donde criadores de Alemania, Inglaterra y Estados Unidos trabajaron simultáneamente, modelando el caótico mosaico original de rayas y manchas en zigzag de los guppies salvajes hasta convertirlo en un diseño simétrico y heredable.
El debate taxonómico y la nomenclatura del patrón
La falta de una nomenclatura científica estandarizada en los inicios de la acuariofilia generó una superposición de términos que aún hoy causa confusión. Conceptos como Lace, Snakeskin, Cobra y King Cobra se han utilizado de manera intercambiable. Sin embargo, esfuerzos de estandarización basados en la morfología del dibujo proponen una clasificación más rigurosa:
- Lace (Encaje): Caracterizado por una red de puntos extremadamente finos y densos.
- Snakeskin (Piel de Serpiente): Presenta un diseño reticulado más grueso, sinuoso y continuo.
- Cobra / Zebrinus (Tigre): Definido por la presencia de barras o rayas verticales conspicuas en la región posterior del pedúnculo caudal.
- King Cobra: Variedad del Snakeskin que destaca por la presencia de manchas melánicas (negras) circulares en los flancos del cuerpo, asemejándose a los "anteojos" característicos de las cobras reales.
Genética de transmisión y retrocruzamiento del Snakeskin
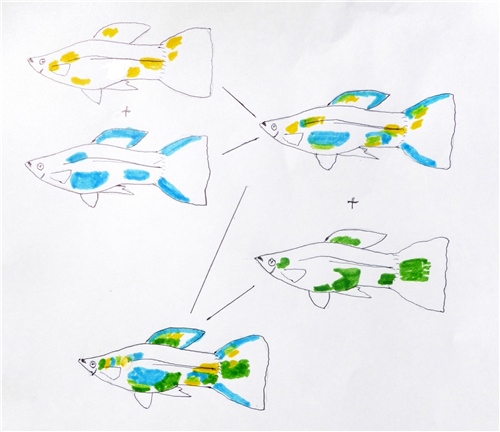





La comprensión de cómo se hereda el patrón Snakeskin requiere analizar los resultados de cruces experimentales controlados. Históricamente, antes de la democratización de la macrofotografía digital de alta velocidad, la ilustración científica detallada desempeñó un papel crucial en el registro de estas transiciones fenotípicas.
[Imagen: Ilustración detallada de la generación F1 obtenida del cruce entre un macho Snakeskin y una hembra multicolor-delta-mosaico]
Al cruzar un macho con patrón Snakeskin corporal y caudal con una hembra de una línea de cuerpo liso y aleta caudal con patrón de mosaico, se observan principios genéticos claros en la primera generación filial ($F_1$):
- Dominancia del patrón corporal: El diseño Snakeskin del cuerpo se comporta de manera dominante frente al cuerpo liso común. No obstante, se evidencia una expresividad variable, ya que el dibujo en la $F_1$ suele presentarse con menor nitidez o definición que en el progenitor puro, sugiriendo la influencia de genes modificadores epistáticos.
- Dominancia del patrón de la aleta caudal: El patrón de la cola tipo "mosaico" muestra dominancia sobre el patrón "lace" (encaje). Los híbridos de la $F_1$ manifiestan la estructura de mosaico, aunque con variaciones en la densidad y gradación del color.
- Jerarquía de pigmentos: Los pigmentos cálidos (amarillos y rojos) muestran una clara dominancia sobre los pigmentos claros o fríos (blancos y azules). La superposición fenotípica del amarillo sobre el azul produce de manera frecuente individuos de coloración verde en las zonas de transición.
El problema de la regresión morfológica
Un fenómeno común al intentar estabilizar el patrón Snakeskin mediante retrocruzamientos (cruzar la descendencia con los progenitores para fijar el carácter) es la pérdida de la estructura de las aletas. Al buscar que el patrón de encaje cubra tanto el cuerpo como la aleta caudal, suele ocurrir una regresión morfológica hacia formas ancestrales de la cola, específicamente hacia la estructura de "doble espada" con bandas.
Esto demuestra que las líneas originales utilizadas para construir el Snakeskin poseían genes estructurales para colas de doble espada. Este tipo de regresión es un recordatorio de que los genes que determinan el color y el patrón a menudo se encuentran estrechamente ligados en los cromosomas a aquellos que determinan la morfología de las aletas.





Fisiología del color: policromía y expresión cromática
Para entender la riqueza visual del guppy, es indispensable abordar la biología de sus células pigmentarias o cromatóforos. La coloración del guppy no es el resultado de un único pigmento, sino de la interacción de varias capas de células especializadas: melanóforos (negro/marrón), eritróforos (rojo), xantóforos (amarillo) e iridóforos (colores estructurales metálicos e iridiscentes).
Dominancia, enmascaramiento y localización
Los pigmentos oscuros y de base química fuerte, como la melanina (negro) y los carotenoides/pteridinas (rojos), tienden a comportarse de forma dominante y tienen la capacidad de enmascarar fenotípicamente a otros colores. Así, un pez puede poseer los genes para expresar azul o verde (genotipo), pero estos pueden permanecer invisibles (fenotipo) si están cubiertos por una capa densa de pigmento rojo o negro.
Además, los cromatóforos no se distribuyen al azar, sino que se organizan en zonas de preferencia o "localizaciones" específicas en el cuerpo del pez.
[Imagen: Esquema de las zonas preferenciales de acumulación de pigmento en el cuerpo del guppy y sus áreas de superposición cromática]
Cuando los colores ocupan solo parcialmente estas zonas, se producen efectos de superposición física y óptica. Por ejemplo, el color verde brillante no siempre se debe a un pigmento verde propiamente dicho, sino a la superposición de células amarillas (xantóforos) sobre una base de células azules estructurales (iridóforos).





El debate entre monocromía y policromía natural
La cría moderna ha tendido a la producción de guppies monocromáticos (azul sólido, rojo sólido, etc.). Si bien esto representa un logro técnico indiscutible debido a la dificultad de suprimir la tendencia natural del pez a la variabilidad, desde una perspectiva biológica y estética se puede argumentar que la monocromía violenta la esencia policromática de la especie. La belleza adaptativa del guppy radica en su capacidad para hacer interactuar armónicamente diferentes colores que destellan según el ángulo de la luz, un fenómeno que se pierde cuando se fija un solo tono plano mediante mutaciones genéticas drásticas (como ocurre en las variedades Japan Blue o Moscow).
La revolución del fenotipo Halfblack o Tuxedo
Hacia 1970, el panorama de la cría de guppies experimentó una de sus mayores transformaciones con la aparición del carácter Halfblack (medio negro), también conocido comercialmente como Tuxedo (esmoquin). Este fenotipo se caracteriza por una coloración negra sólida y densa que cubre la mitad posterior del cuerpo del pez, creando un contraste radical con la mitad anterior y las aletas.
Un origen envuelto en la niebla genética
Aunque los guppies silvestres suelen presentar pequeñas manchas o barras de melanina en el pedúnculo caudal, el mecanismo por el cual estas pequeñas "islas" de pigmento se fusionaron para formar una capa negra continua y uniforme sigue siendo un misterio evolutivo. La asignación del término científico nigrocaudatus para describir esta variedad simplemente catalogó el fenómeno sin explicar la ruta mutacional que lo hizo posible.
Desde el punto de vista genético, el carácter Halfblack presenta particularidades muy marcadas:
- Superdominancia: Una vez introducido en una línea de cría, es extremadamente difícil de eliminar, comportándose de manera dominante sobre casi cualquier otra distribución de melanóforos.
- Herencia no ligada exclusivamente al sexo: A diferencia de muchos otros patrones de color en el guppy que se transmiten exclusivamente a través del cromosoma Y (de padres a hijos machos), el factor Halfblack se expresa tanto en machos como en hembras, lo que indica una herencia autosómica o una translocación cromosómica con alta penetrancia en ambos sexos.





Estética y salud biológica: el peligro de la hiper-selección
El fenotipo Tuxedo ofrece un excelente lienzo debido al contraste que genera. Los ejemplares más valiosos desde el punto de vista estético son aquellos donde la monotonía del negro se rompe en la parte anterior del cuerpo mediante destellos metálicos, irisaciones fluorescentes en la aleta dorsal y un contraste limpio.
[Imagen: Ejemplar de guppy Halfblack que muestra la raya dorsal fluorescente en contraste con el cuerpo negro sólido]
Sin embargo, la selección extrema de este y otros caracteres ha llevado en la actualidad al desarrollo de variedades morfológicamente hipertrofiadas, como las aletas pectorales gigantes (Dumbo) o las aletas caudales sobredimensionadas (Full Moon). Desde la perspectiva de la morfología funcional y la hidrodinámica, estas características resultan perjudiciales para el bienestar del pez, pues limitan su capacidad natural de natación y aumentan su susceptibilidad a enfermedades físicas, transformando un organismo biológicamente eficiente en un fenotipo meramente ornamental e inviable en condiciones naturales.
Conclusiones
La historia evolutiva y la cría selectiva del guppy demuestran que la genética de esta especie es un sistema dinámico y multifactorial que va más allá de las leyes de la dominancia simple de Mendel. Variedades emblemáticas como el Snakeskin y el Tuxedo ilustran cómo la selección artificial puede moldear caracteres latentes en las poblaciones silvestres hasta crear formas de una complejidad asombrosa. No obstante, el éxito de la cría de conservación y de exposición depende del respeto a la fisiología natural del pez. El verdadero arte de la cría selectiva no consiste en forzar al organismo hacia la monotonía estética o la deformidad funcional, sino en comprender y potenciar los mecanismos heredados de policromía e iridiscencia que hacen del guppy un auténtico lienzo viviente de la evolución.