14. El lienzo genético del guppy: Complejidad, mutación y patrones en la acuariofilia de precisión
De las leyes mendelianas a la teoría de segmentación corporal en la expresión fenotípica de Poecilia reticulata
Autor: Javier Rodríguez Luna
Fecha: 29 de Mayo del año 2026
Introducción
El guppy (Poecilia reticulata) ha sido, durante más de un siglo, uno de los organismos modelo más fascinantes tanto para la investigación biológica como para la acuariofilia de selección. Su alta tasa reproductiva y su asombrosa plasticidad fenotípica ofrecen un laboratorio vivo para el estudio de la transmisión de caracteres. Sin embargo, lo que a menudo se presenta como un simple ejercicio de genética mendeliana revela, bajo un análisis más riguroso, una red de interacciones poligénicas, mutaciones espontáneas y patrones de distribución espacial que desafían los modelos más simplistas. Este artículo analiza cómo las leyes de la herencia interactúan con la complejidad molecular, los mecanismos de mutación como el crossing-over y la revolucionaria propuesta de la segmentación corporal en cuadrícula para descifrar la infinita variabilidad cromática de esta especie.


#GeneticaAcuatica #DivulgacionCientifica #Acuariofilia #PoeciliaReticulata #EvolucionYGenetica
Más allá de Mendel: La complejidad de la herencia poligénica
El salto cuantitativo: De un par de genes a múltiples alelos
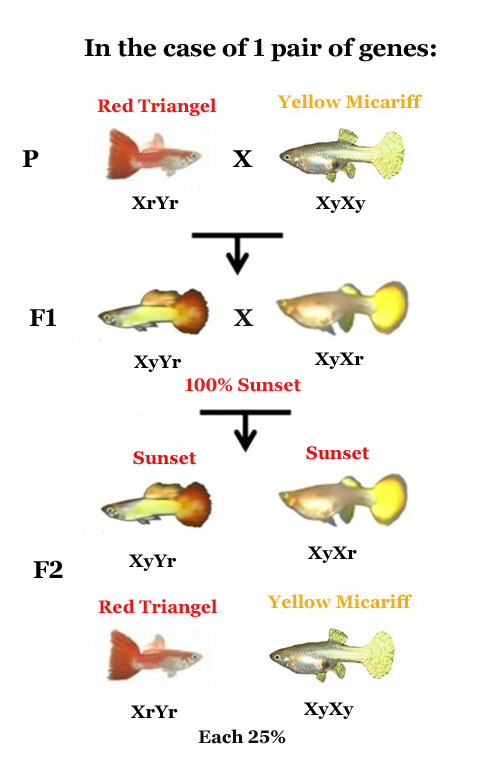
La genética clásica suele introducirse mediante el cruce de caracteres determinados por un único par de genes con alelos dominantes y recesivos. En este escenario simplificado, el cruce de homocigotos produce una primera generación filial (F1) uniforme de heterocigotos que expresan el fenotipo dominante. Al cruzar esta F1 entre sí, la segunda generación filial (F2) arroja una proporción predecible de cuatro combinaciones genotípicas básicas, donde el carácter recesivo reaparece en un 25% de los casos.
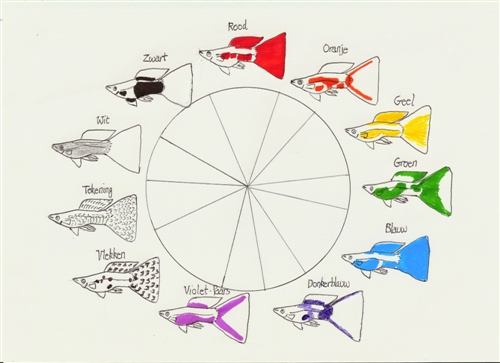




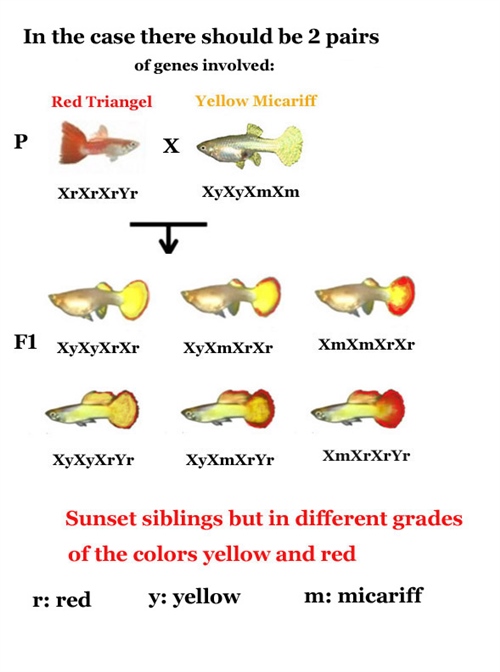
Sin embargo, la realidad fenotípica del guppy es multifactorial. Cuando en la determinación de un carácter interviene más de un par de genes (multiplicidad de alelos y loci), la complejidad matemática y biológica se expande exponencialmente. En un cruce que involucra dos pares de alelos, el número de combinaciones posibles en la descendencia asciende a 16, dando lugar a una variedad de genotipos y fenotipos intermedios que reflejan con mayor fidelidad la diversidad observable en los criaderos modernos.
[Imagen: Esquema comparativo entre un cruce monohíbrido clásico y un cruce dihíbrido poligénico]
Calidad contra cantidad: El gradiente cromático
Un aspecto crítico en la herencia poligénica es que un carácter no solo está determinado por la presencia de un gen específico (un efecto cualitativo), sino también por la dosis o cantidad de genes acumulados (un efecto cuantitativo).





Un ejemplo paradigmático es la coloración de la aleta caudal conocida como "Sunset" (Atardecer). En este caso, la intensidad de la coloración roja o amarilla no se comporta como un interruptor de "encendido o apagado". Cuantos más genes promotores del rojo estén presentes en el genoma del pez, más intensa y dominante será dicha tonalidad en la aleta. Por el contrario, a mayor densidad de genes para el amarillo, más clara y dorada será la aleta. Si se introduce un carácter recesivo, como el amarillo "Micariff", este no logrará expresarse en la F1 debido a la dominancia de otros alelos, pero reaparecerá en la F2 en una proporción matemática significativamente menor que la observada en los cruces de un solo gen.
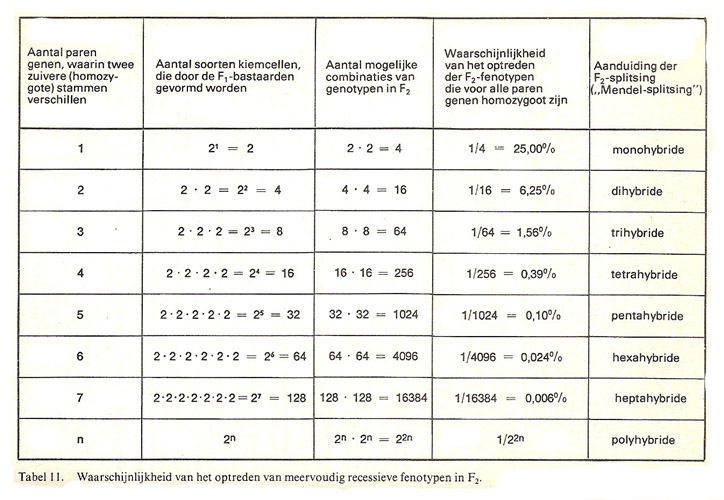
El cálculo probabilístico de la rareza
La probabilidad de obtener fenotipos específicos disminuye drásticamente a medida que aumenta el número de loci génicos involucrados. La aparición de caracteres recesivos múltiples sigue una curva de probabilidad decreciente:
- Un carácter recesivo simple: se expresa en el 25% de la descendencia F2 (1 de cada 4).
- Tres caracteres recesivos independientes (como la condición triple recesiva del guppy albino): se expresa en apenas el 1,56% de los individuos.
- Cinco pares de alelos: la probabilidad se reduce al 0,1%.
- Siete pares de alelos: el fenotipo deseado se convierte en una rareza de 1 entre 16 500 ejemplares.






Si proyectamos este cálculo a un escenario real donde se analicen, por ejemplo, 20 loci génicos para determinar 10 colores posibles, las combinaciones genotípicas potenciales superan los 10 millones. Si a esto añadimos los patrones de dibujo y las diversas formas de la aleta caudal (espada, delta, lira, etc.), nos enfrentamos a una cifra astronómica de variaciones posibles. Esto explica por qué la fijación de líneas puras de exhibición requiere un control riguroso de la endogamia y una selección fenotípica meticulosa.
El enigma del Micariff: Mutaciones y flujo genético en la creación de variedades
Un misterio de Sri Lanka




La historia de la variedad "Micariff" ilustra a la perfección cómo el desarrollo de nuevas líneas combina el misterio comercial con los mecanismos evolutivos de la genética de poblaciones. Surgido hace aproximadamente un cuarto de siglo en Sri Lanka, el Micariff destaca por poseer un color amarillo metálico claro, de un brillo distintivo que lo diferencia notablemente del clásico "German Yellow" (Amarillo Alemán).
Durante años, debido al secretismo de los criadores comerciales, se especuló sobre su origen. Algunas hipótesis sugerían que era el resultado de hibridar la variedad metálica "Stoerzbach" con el patrón "Snakeskin" (Piel de Serpiente) amarillo. No obstante, las diferencias morfológicas y de distribución pigmentaria sembraron dudas: los Stoerzbach presentan una pigmentación irregular y rugosa, mientras que el Micariff exhibe una distribución fina, homogénea y brillante. Otras teorías propusieron cruces con "Yellow Full Platinum", "Full Gold" o incluso variantes invertidas de "Half Black". El hecho de que el Micariff sea genéticamente compatible con la línea "Moscow" para introducir el pigmento amarillo refuerza la idea de que nos encontramos ante una pieza clave del rompecabezas genético de la especie.
[Imagen: Comparativa fenotípica entre variedades de guppy: Micariff, Stoerzbach y Yellow Full Platinum]
El mecanismo del crossing-over como motor de la diversidad
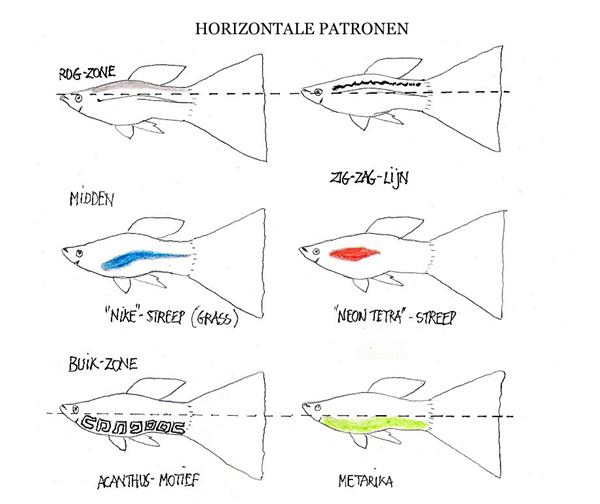
Para evitar debates interminables sobre cruces hipotéticos, la biología evolutiva ofrece una explicación más elegante: la mutación y la recombinación meiótica. Durante la formación de los gametos (óvulos y espermatozoides), los cromosomas homólogos se aparean y se enrollan estrechamente. En este proceso, denominado crossing-over (entrecruzamiento), las hebras de ADN pueden sufrir roturas y uniones recíprocas accidentales.
Este intercambio de fragmentos a nivel cromosómico profundo rompe los ligamientos génicos preexistentes, reordenando los alelos y dando lugar a combinaciones genéticas inéditas. Cuando este proceso ocurre de forma atípica o se ve influenciado por factores externos (como la radiación ultravioleta o condiciones severas de estrés ambiental), pueden surgir mutaciones estables que alteran el fenotipo, dando origen a variedades únicas como el Micariff.
La búsqueda de un orden: De los mapas clásicos a la teoría de la segmentación corporal
Los pioneros y el desafío de la sobreinformación
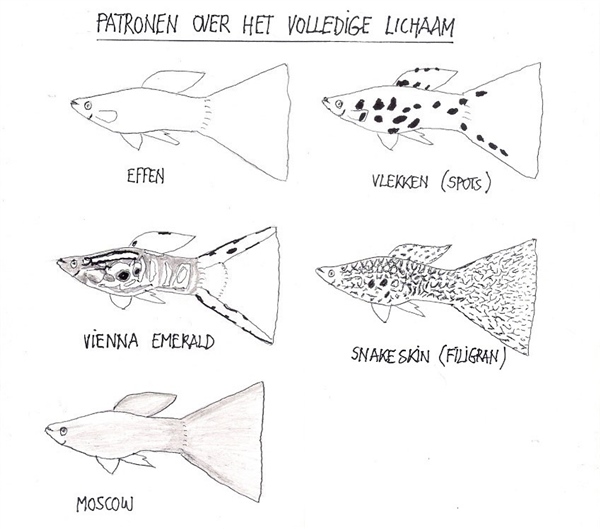
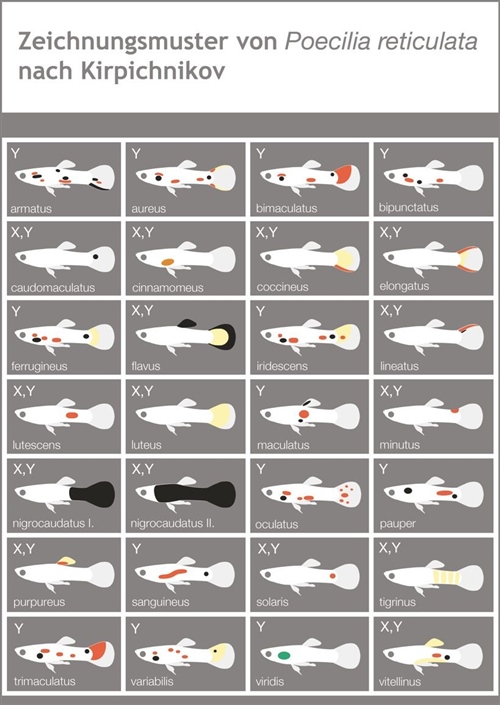
A mediados del siglo XX, genetistas pioneros como el danés O. Winge y el ruso V. Kirpichnikov dedicaron décadas a cartografiar y fijar los patrones del guppy. A pesar del valor histórico de sus trabajos, sus aproximaciones adolecieron de un exceso de especificidad anecdótica. Al intentar registrar de forma individualizada cada variante de color en zonas hiperespecíficas del cuerpo del pez (como el tipo cinnamomeus o canela de Winge), perdieron de vista las líneas generales de organización genómica. Kirpichnikov avanzó sustancialmente, pero cayó en el mismo error de sobrecargar sus mapas con detalles minuciosos, logrando definir con absoluta claridad únicamente el patrón nigrocaudatus (cola negra).
El triple funcionamiento génico: Tipo, intensidad y localización
Para obtener una visión de conjunto coherente y aplicable a la cría práctica, se propone el principio del triple funcionamiento de los genes. Este modelo postula que la expresión de los caracteres pigmentarios en el guppy se rige por tres variables interconectadas:
- Calidad (Tipo): Determina el color o patrón específico (por ejemplo, pigmento negro, rojo o amarillo).
- Cantidad (Intensidad): Regula la densidad y saturación de dicho pigmento según la carga alélica presente.
- Ubicación (Localización corporal): Define el área anatómica precisa donde se activarán o silenciarán los genes responsables de dicho pigmento.
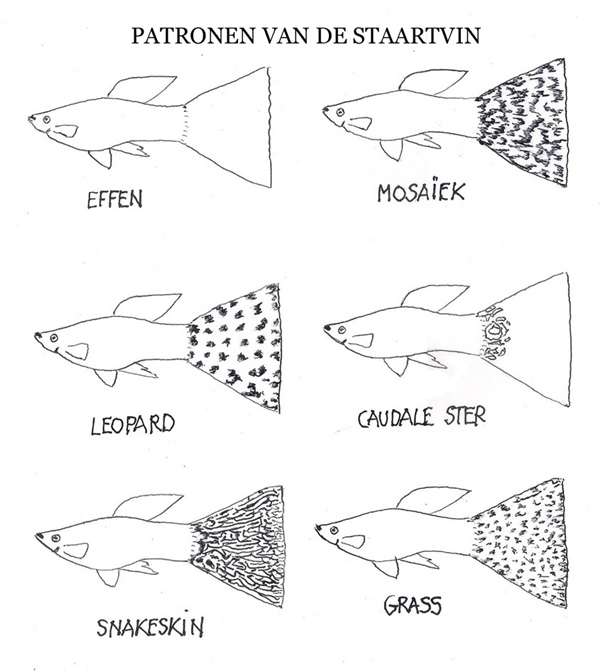
En el mapa de dominancia general, el pigmento negro ejerce una dominancia casi absoluta sobre el resto de los colores. El rojo ocupa el segundo lugar, pero posee una notable capacidad para crear coloraciones mixtas en las zonas de solapamiento (dando origen a tonos naranjas, marrones, púrpuras o rosados). Por su parte, el verde y el azul se comportan de manera equivalente, fusionándose en un tono azul verdoso allí donde coinciden.
El modelo 3x3: Segmentación y los bloques fijos del genoma
La cuadrícula corporal del guppy
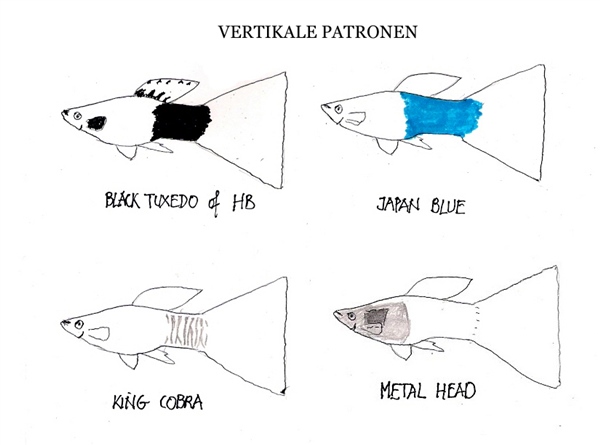
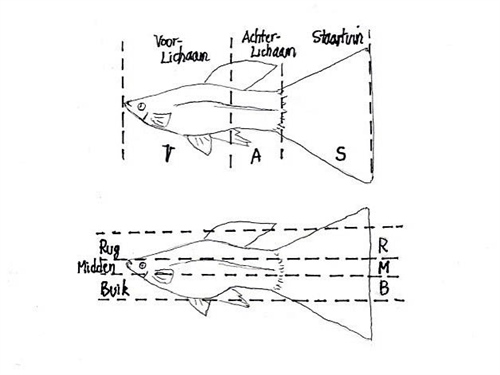
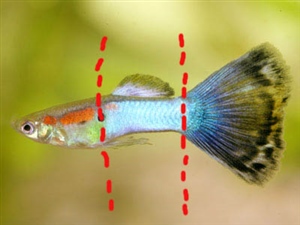
Una de las aportaciones teóricas más relevantes para comprender la variabilidad fenotípica sin perderse en el caos de los detalles es la hipótesis de la segmentación en cuadrícula. Según este modelo, el cuerpo del guppy se divide funcionalmente en tres segmentos horizontales y tres segmentos verticales, configurando una matriz de nueve zonas de desarrollo independiente.
[Imagen: Diagrama del cuerpo de un guppy dividido en una matriz de 3x3 segmentos horizontales y verticales]
Esta segmentación anatómica explica dos fenómenos cruciales para la genética aplicada:
- La formación de bloques genómicos estables: Determinados fragmentos del genoma se agrupan estrechamente para constituir patrones fenotípicos fijos y altamente resistentes a la separación. Estos bloques se comportan de manera "bloqueada", impidiendo que otros alelos ocupen su lugar y asegurando la estabilidad de la línea. Se asemeja a una ruta neuronal o a un hábito biológico: la maquinaria celular tiende a replicar el camino genético más transitado.
- La deconstrucción controlada de patrones: A pesar de su estabilidad, estos bloques no son herméticos. Bajo procesos selectivos o de hibridación, los patrones pueden desensamblarse siguiendo precisamente las líneas de fractura de los segmentos horizontales y verticales, lo que permite a los criadores aislar o desplazar rasgos cromáticos a zonas específicas del cuerpo del pez.
Las siete columnas de la naturaleza
Aunque en los acuarios de selección encontramos una aparente infinitud de formas y colores, la genética de poblaciones silvestres nos recuerda que la naturaleza es notablemente económica. En el medio natural, la inmensa mayoría de las poblaciones de guppys se organizan en torno a únicamente siete patrones básicos de herencia, a partir de los cuales la selección artificial humana ha esculpido todas las variedades modernas durante el último siglo:
- Sólido (Liso): Coloración uniforme en todo el cuerpo.
- Cola Mosaico: Patrón de manchas oscuras entrelazadas sobre fondo claro en la caudal.
- Pauper (Marmolado): Distribución irregular y moteada de pigmentos.
- Estrella Caudal: Concentración pigmentaria en el centro o base de la aleta caudal.
- Cola de Encaje (Lace): Filigrana fina y reticulada de aspecto textil.
- Nigrocaudatus: Melanización intensa de la mitad posterior del cuerpo.
- Ojo de Pájaro: Dibujo circular concéntrico con alto contraste.
Conclusiones
La crianza y selección del guppy de fantasía trasciende la mera afición para consolidarse como un ejercicio de genética aplicada de alta precisión. La transición desde los modelos monogénicos simples hacia la comprensión de la herencia poligénica, las dinámicas de mutación por crossing-over y la revolucionaria teoría de la segmentación corporal en matriz de 3x3, ofrece una estructura científica sólida para predecir y fijar caracteres. Comprender que cada línea de guppys no es una entidad estática aislada, sino una "instantánea" temporal dentro del flujo continuo de un acervo génico dinámico, permite a los criadores trabajar en armonía con las leyes orgánicas de la naturaleza, transformando el acuario en un lienzo donde la ciencia y la estética convergen de manera perfecta.